
イントロダクション
中生代恐竜類における熱生理学的多様性と代謝進化:地球化学および分子古生物学的アプローチによる最新知見の統合報告
恐竜の熱生理学、すなわち彼らが「温血(内温性)」であったか「冷血(外温性)」であったかという問いは、1842年にリチャード・オーウェンが「Dinosauria」を定義して以来、古生物学における中心的な論争の主題となってきました。19世紀の鈍重な爬虫類というイメージから、1960年代から70年代の「恐竜ルネサンス」を経て鳥類に近い活動的な動物像への転換、そして21世紀における直接的な生理指標の測定技術の確立に至るまで、我々の恐竜観は劇的な変遷を遂げてきました。近年の研究は、恐竜が単一の代謝戦略を採用していたのではなく、系統ごとに極めて多様で動的な熱生理学的進化を遂げていたことを明らかにしています。
恐竜の代謝と体温調節:定義と研究史の背景
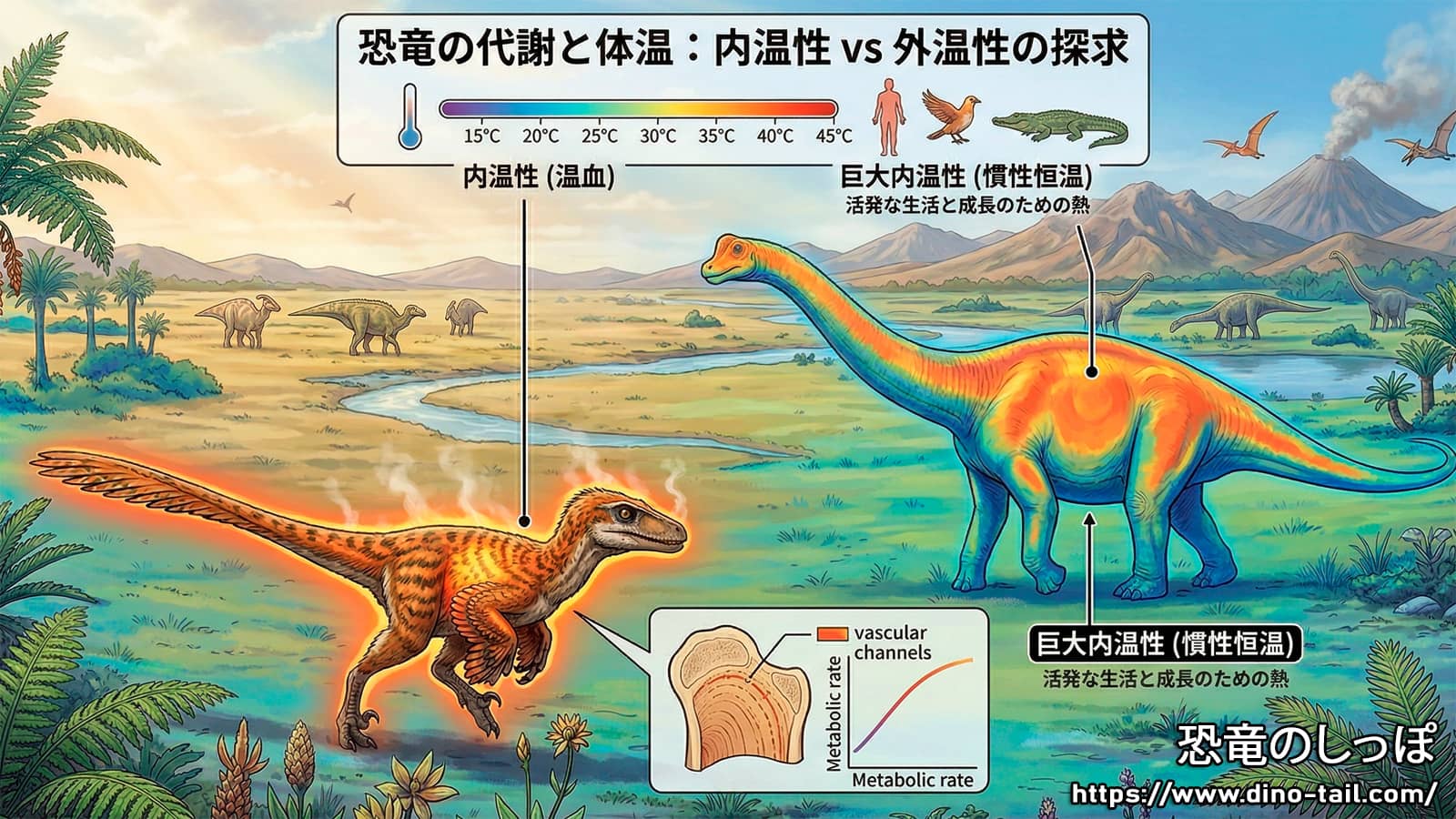
恐竜の熱生理を理解するためには、まず生物学における代謝と体温調節の概念を整理する必要があります。現代の動物は、代謝によって内部で熱を生成する内温動物(哺乳類・鳥類)と、外部環境からの熱に依存する外温動物(爬虫類・両生類・魚類)に大別されます。内温動物は高い代謝率(BMR)を持ち、安定した高い体温(恒温性)を維持することで、環境に左右されない高い活動性を確保していますが、その代償として大量の食料を必要とします。
恐竜の研究史において、初期の推定は主に骨組織学(骨の成長速度)や解剖学的特徴(直立歩行の姿勢)に基づいていました。しかし、これらの指標は間接的なものに過ぎず、成長速度が速いからといって必ずしも現代の鳥類のような高度な内温性を備えていたとは限らないという反論も根強くありました。2014年には、成長速度の全学的解析に基づき、恐竜が内温性と外温性の中間に位置する「中温性(Mesothermy)」であったとする説が提唱されましたが、このモデルもスケーリング手法の妥当性を巡って激しい議論を呼んでいます。
塊状同位体温度計による直接的体温推定
21世紀に入り、地球化学的アプローチの一環として「塊状同位体温度計(Clumped Isotope Paleothermometry)」が導入されたことで、化石から直接的に体内温度を算出することが可能となりました。この手法は、炭酸塩鉱物(歯のエナメル質や卵殻)の結晶格子内において、重い同位体である炭素13(13C)と酸素18(18O)が結合(クランプ)する割合が、鉱物形成時の温度に依存するという物理化学的原理に基づいています。従来の酸素同位体比(δ18O)を用いた分析とは異なり、周囲の環境水の同位体組成を知る必要がないため、極めて信頼性の高い「古温度計」として機能します。
卵殻および歯の同位体分析による分類群別データ
カリフォルニア工科大学、イェール大学、UCLAなどの研究チームが行った一連の分析により、主要な恐竜グループの体内温度が明らかにされてきました。以下の表は、塊状同位体(Δ47)に基づいた主要な恐竜の体内温度推定値と、当時の環境推定温度を比較したものです。
| 恐竜の分類群 | 推定体内温度(℃) | 環境推定温度(℃) | 備考 |
|---|---|---|---|
| ブラキオサウルス ( Brachiosaurus ) | 38.2(℃) | 25(℃) | 巨大竜脚類。高い体温を維持。 |
| カマラサウルス ( Camarasaurus ) | 35.7(℃) | 25(℃) | 竜脚類。哺乳類に近い温度。 |
| マイヤサウラ ( Maiasaura ) | 44(℃) | 25(℃) | 鳥盤類。極めて高い内温性を示唆。 |
| トロオドン ( Troodon ) | 27, 28(℃) | 25~(℃) | 獣脚類。不均一な体温。 |
| オヴィラプトル類 ( Oviraptorid ) | 32(℃) | 26(℃) | 小型獣脚類。環境よりわずかに高い。 |
| チタノサウルス類(矮小種) | 36(℃) | 30(℃) | 小型化しても高温を維持。代謝制御の証拠。 |
これらのデータから導き出される重要な洞察は、恐竜が体サイズに関わらず、代謝によって体温を環境温度以上に引き上げる能力を共通して持っていたという点です。特に、比較的小型のトロオドンや中型のマイヤサウラが環境温度を大きく上回る体温を示していることは、これらが単なる熱慣性(巨大な体が冷めにくい現象)に頼る外温動物ではなく、能動的な熱生成を行っていたことを裏付けています。
二重塊状同位体(Δ47 と Δ48)による高度な検証
最新の研究では、Δ47 に加えて Δ48を測定する「二重塊状同位体温度計」が導入され、鉱物形成プロセスの非平衡性(キネティック効果)までもが考慮されるようになりました。トロオドンの卵殻を用いた分析では、鳥類に見られる高度な非平衡プロセスとは異なり、爬虫類に近い平衡状態での鉱物形成が確認されました。しかし、体内温度自体は環境よりも高く、かつ個体間や生理状態で変動する「異温性(Heterothermy)」を伴う内温性であったことが示唆されています。これは、恐竜から鳥類への進化過程において、恒温性が段階的に獲得されていったことを示す重要な証拠です。
分子古生物学の革新:代謝マーカーによる直接測定
2022年に『Nature』誌に掲載されたヤスミナ・ヴィーマンらの研究は、恐竜の熱生理学研究を根底から覆す革新的な手法を提示しました。この手法は、骨組織内の同位体を測るのではなく、動物が呼吸によって取り込んだ酸素が代謝される際に発生する「化学的老廃物」を直接検出するものです。
高度な脂質酸化終末産物(ALEs)の検出
細胞内の代謝プロセスにおいて酸素が消費されると、活性酸素種(ROS)が副産物として生成されます。これが脂質、タンパク質、糖と反応し、硬組織内に蓄積されるのが「高度な脂質酸化終末産物(ALEs)」や「高度な糖化終末産物(AGEs)」です。これらの分子は極めて安定しており、化石化の過程でも失われず、その蓄積量は動物が生前に消費した酸素量、すなわち代謝率と正の相関を示します。
ヴィーマンらは、ラマン分光法およびフーリエ変換赤外分光法(FT-IR)を用いて、55種に及ぶ現代および絶滅した脊椎動物の骨(主に大腿骨)を分析しました。この非破壊的な手法により、恐竜の系統樹全体における代謝率の分布が初めて定量化されました。
竜盤類と鳥盤類の代謝的分離
分析の結果、恐竜の代謝戦略は主要な系統間で驚くべきコントラストを見せることが判明しました。
- 獣脚類(Theropoda) : ティラノサウルスやヴェロキラプトル、そして現代の鳥類を含むこのグループは、極めて高い代謝率を示しました。一部の種では現代の鳥類を凌駕する代謝率を持っており、これらが「真の内温動物」として活動的な捕食者であったことが確認されました。
- 竜脚類(Sauropodomorpha) : ブラキオサウルスやディプロドクスなどの巨大な草食恐竜もまた、内温動物に匹敵する高い代謝率を維持していました。これは、彼らの高温の維持が単なる巨大な体躯による熱慣性(ギガントサーミー)によるものだとする従来の有力な仮説を否定し、能動的な生理機能によるものであることを示しています。
- 鳥盤類(Ornithischia) : トリケラトプス、ステゴサウルス、ハドロサウルス類を含むこのグループは、対照的に極めて低い代謝率を示しました。その数値は現代のトカゲやカメと同等であり、これらは二次的に外温性へと進化したか、あるいは低い代謝率で環境に適応していた可能性が浮き彫りになりました。
この知見は、恐竜が共通の祖先から高代謝を引き継いだものの、鳥盤類においてはこの特性が失われたことを示唆しており、恐竜の進化が単純な「温血化」の道筋を辿ったのではないことを物語っています。
進化的トリガーと気候適応:内温性の起源
恐竜がいつ、なぜ内温性を獲得したのかという問題に対し、2024年にアルフィオ・アレッサンドロ・キアレンツァらが発表した研究は、化石の地理的分布と古気候モデルを組み合わせることで新たな視点を提供しています。
ジュラ紀初期の気候変動とジェンキンス・イベント
約1億8300万年前、ジュラ紀初期に発生した「ジェンキンス・イベント」は、大規模な火山活動に伴う急激な地球温暖化と植物相の変化をもたらしました。この環境ストレスが、恐竜の系統間で異なる生理学的進化を促したと考えられています。
研究チームは1000点以上の化石記録を分析し、獣脚類と鳥盤類がこの時期に、より寒冷な高緯度地域へと進出していったことを突き止めました。一方で、竜脚類は温暖な低緯度地域に留まり続けました。この分布パターンの変化は、獣脚類と鳥盤類が寒冷地での活動を可能にする内温性(および断熱のための羽毛)を発達させた時期と一致します。
進化的帰結:適応と制約
内温性の獲得は、恐竜に高い成長速度と、寒冷地を含む多様な環境への進出という大きなアドバンテージを与えました。しかし、巨大な体躯を持つ竜脚類にとっては、高い基礎代謝は放熱を困難にし、過熱のリスクを高める制約となった可能性があります。竜脚類が熱帯・亜熱帯地域に限定されていた事実は、彼らが環境温度を巧みに利用する外温的な生理、あるいは制御された「熱慣性(ギガントサーミー)」に頼っていたことを示唆しています。
研究間の矛盾と論争の解析:マイヤサウラの事例
最新の研究成果(ヴィーマン 2022)と、それ以前の同位体研究(ドーソン 2020)の間には、鳥盤類の代謝を巡る顕著な不一致が存在します。
矛盾の核心
- 同位体データ(ドーソン 2020) : 鳥盤類のマイヤサウラは44℃という極めて高い体内温度を示し、高度な代謝制御を行っていたとされます。
- 分子マーカーデータ(ヴィーマン 2022) : ハドロサウルス類(マイヤサウラの仲間)はトカゲ並みの低代謝率であると報告されています。
この矛盾を解釈する鍵は、代謝率と体内温度の「分離」にあります。マイヤサウラのような中型から大型の恐竜は、たとえ基礎代謝率が低くても、熱慣性によって高い体温を維持することが可能です。また、特定の活動時や繁殖期にのみ代謝を上げる通性的内温性(Facultative Endothermy)を持っていた可能性も考えられます。さらに、骨の形成部位(末端か中枢か)によって温度や代謝産物の蓄積量が異なる「熱モザイク」の可能性も、今後検証すべき課題です。
生理機能と形態的証拠の相関:脳、心臓、そして呼吸
代謝率の高さは、その動物の活動レベルだけでなく、解剖学的な複雑さとも密接に関連しています。
循環器系と呼吸器系の進化
高い代謝を維持するためには、組織への効率的な酸素供給が不可欠です。
- 心臓の構造 : 高い代謝率を持つ獣脚類や竜脚類は、哺乳類や鳥類と同様に、肺循環と体循環を完全に分離した強力な四室心臓を持っていたと推測されます。
- 気嚢システム : 獣脚類や竜脚類の骨に見られる空洞(気腔)は、鳥類と同様の一方向性呼吸システム(気嚢システム)の存在を示唆しています。これは呼吸効率を最大化するだけでなく、巨大な体内からの放熱を助けるクーラーとしても機能したと考えられます。
知能と脳のサイズ
近年の神経解剖学的な研究では、代謝率と脳のサイズ(脳化指数)の相関が議論されています。
- 獣脚類 : ティラノサウルスなどの大型獣脚類は、高い代謝率に支えられ、霊長類に近い神経細胞密度を持っていた可能性が指摘されています。
- 鳥盤類と竜脚類 : これらは対照的に相対的な脳サイズが小さく、これは彼らの低代謝(特に鳥盤類)や、エネルギーを身体の維持に特化させた戦略と一致します。
DAEDALUS プロジェクト:内耳形態による新しいアプローチ
現在進行中の「DAEDALUS プロジェクト」は、化学的保存状態に左右されない新しい体温推定手法を開発しています。
脊椎動物の内耳にある半規管は、平衡感覚を司る器官です。この中を満たす内リンパ液の粘性は、温度によって劇的に変化します。
熱可動性指数(TMI) : リカルド・アラウージョらのチームは、半規管の形状とサイズの比率から、その動物が最も効率的に平衡感覚を機能させられる「設計温度」を算出するモデルを構築しました。
AIとナノ粘度計の導入 : 最新のAIアルゴリズムとナノレベルの粘度測定技術を用いることで、化石のCTスキャンデータから体内温度を逆算するこの試みは、化学分析(同位体や分子マーカー)を補完する第三の柱として期待されています。
まとめ:中生代における熱生理学のランドスケープ
恐竜の熱生理学に関する最新の研究成果は、従来の単純な二元論を脱却し、複数の要素が重なる多角的な分析が必要なことを示しています。
- 内温性の広範な分布 : 塊状同位体分析は、恐竜の主要な全系統において、環境温度を上回る代謝制御が存在したことを示しています。代謝熱の生成能力は、恐竜の祖先において早期に進化した形質でした。
- 系統間の代謝的分化 : 分子マーカーによる分析は、内温性の強度が系統によって大きく異なっていたことを明らかにしました。獣脚類と竜脚類が「高代謝・活動的」な戦略を維持したのに対し、鳥盤類は「低代謝・省エネ」な戦略へとシフトした可能性が高いです。
- 環境との相互作用 : ジュラ紀初期の気候変動が、恐竜の生理的適応を加速させ、それによって彼らは高緯度から熱帯まで地球全体を支配するに至りました。
- 活動性と適応戦略 : 高い代謝は、羽毛の発達、巨大な体躯の維持、高度な知能の獲得、そして最終的には鳥類による「飛行」の実現を可能にしました。
恐竜の体温調節に関する知見は、単に過去の動物の温度を測るだけでなく、生物がどのようにしてエネルギーの限界を突破し、地球規模の支配者へと進化していったのかという、生命の根本的な戦略を解き明かす鍵となっています。今後、DAEDALUSプロジェクトに代表される新しい物理的アプローチが加わることで、恐竜の熱生理学はより精密で統合的な理解へと向かうでしょう。
| 分類群 | 代謝率の推定 | 体内温度の安定性 | 主な適応・特徴 |
|---|---|---|---|
| 獣脚類 | 非常に高い(鳥類級) | 恒温 または 異温 | 羽毛、気嚢、高い知能、捕食活動、飛行の起源。 |
| 竜脚類 | 高い(哺乳類級) | 恒温(熱慣性併用) | 巨大な体、長い首による放熱、高効率な消化。 |
| 鳥盤類 (ハドロサウルス類) | 低い(爬虫類級)〜 中程度 | 変温 または 恒温(熱慣性) | 複雑な歯、群れ行動、一部で二次的な低代謝化。 |
| 鳥盤類 (装盾類・周飾頭類) | 低い | 変温に近い | 鎧や角、低い活動レベル、環境依存の体温調節。 |
| 翼竜 | 高い | 恒温 | 飛行に必要な高エネルギー代謝、恐竜と共通の祖先。 |